Volumen 31
Número 60
Mayo - Octubre 2020
ISSN: 1851-1716
Volumen 31
Número 60
Mayo - Octubre 2020
ISSN: 1851-1716
Ciencias Exactas y Naturales – Investigación
Análisis de polimorfismos en los genes SOX6 y Ryr1 y su relación con la calidad de carne de cerdo
Genetic polymorphisms analysis of Ryr1 and SOX6 and porcine meat quality traits
Análise de polimorfismos nos genes SOX6 e Ryr1 e sua relação com a qualidade da carne suína
Análisis de polimorfismos en los genes SOX6 y Ryr1 y su relación con la calidad de carne de cerdo
Ciencia, Docencia y Tecnología, vol. 31, núm. 60, 2020
Universidad Nacional de Entre Ríos
Recepción: 05 Abril 2019
Aprobación: 06 Marzo 2020
Resumen: Se estudió la incidencia de polimorfismos de los genes Ryr1 y SOX6 en una población porcina del noreste entrerriano. Ambos genes están relacionados con la calidad de carne, Ryr1 es responsable del síndrome de estrés porcino y carnes pse, mientras que SOX6 es un factor de transcripción importante en la diferenciación de fibras musculares. Se analizó el efecto de dos SNPs de SOX6 y uno de Ryr1 con respecto a los atributos de calidad. Los resultados mostraron que el pH y la cra se vieron influenciados significativamente con SOX6a mientras que SOX6b solo se vinculó con el pH. La implementación de este tipo de estrategias contribuye al crecimiento de la industria porcina y a responder a las exigencias de los consumidores. Este trabajo sugiere que la selección de genotipos portadores de alelos C para Ryr1, A para SOX6a y C para SOX6b podría mejorar la producción de carne fresca de buena calidad.
Palabras clave: Calidad de carne, Cerdo, Halotano, SOX6, pH, cra.
Abstract: The incidence of Ryr1 and SOX6 polymorphisms were studied in a population of northeastern Entre Ríos. Both genes are related to meat quality, Hal is known to be responsible for porcine stress syndrome and the presence of pale, soft and exudative meat (pse) while SOX6 is a versatile transcription factor and plays an important role in muscle fiber differentiation. The markers at SOX6 loci were tested for their interactions/effect with Ryr1 regarding quality traits. Results showed that the pH and cra were significantly related to SOX6a while only the ph of the meat was related with SOX6b. Strategy formulation based on this approach will help to support a growing pork industry and respond to consumer demand. Therefore, this work suggests that the selection of genotypes carrying C alleles for Halothane, A for SOX6a and C for SOX6b could improve the production capacity of fresh meat of good quality.
Keywords: Meat quality, Pork, Halotane, SOX6, pH, whc.
Resumo: Foi estudada a incidência de polimorfismos dos genes Ryr1 e SOX6 em uma população de suínos do nordeste de Entre Rios. Ambos os genes estão relacionados com a qualidade da carne, Ryr1 é responsável pela síndrome de estresse porcino e carnes pse, enquanto o SOX6 é um importante fator de transcrição na diferenciação das fibras musculares. Foi analisado o efeito de dois SNPs de SOX6 e um de Ryr1 em relação aos atributos de qualidade. Os resultados mostraram que o pH e a cra foram significativamente influenciados pelo SOX6a, enquanto o SOX6b foi associado apenas ao pH. A implementação deste tipo de estratégias contribui para o crescimento da indústria de suínos e para responder às demandas dos consumidores. Este trabalho sugere que a seleção dos genótipos portadores de alelos C para Ryr1, A para SOX6a e C para SOX6b poderia melhorar a produção de carne fresca de boa qualidade.
Palavras-chave: Qualidade de carne, Porco, Halotano, SOX6, pH, CRA.
Introducción
Dada la importancia de la industria de la carne porcina y la exigente demanda de los consumidores, las estrategias que favorecen una mejora en su calidad se encuentran en un proceso de desarrollo continuo. La percepción de la calidad de carne depende de un concepto subjetivo del consumidor, quien ya no solo está exigiendo un alto contenido magro de la canal sino también terneza, marmolado, aroma y acidez óptimos en adición a las características que la hacen elegible en góndola como el color y la capacidad de retención de agua (cra). El concepto calidad de carne está relacionado además a componentes nutricionales, higiénicos, tecnológicos y genéticos como así también a factores del metabolismo celular que influyen en los atributos de la carne (Bastos et al., 2001, Marini et al., 2012). Por lo tanto, ante las mayores exigencias del mercado, la producción de la carne de cerdo debe abarcar todos los puntos que constituyen la cadena de producción (Chan et al., 1997; Chan et al., 1998; Faustman et al., 1998; Suman et al., 2007, Vestergaard et al., 2000). La carga genética de los animales es un factor fundamental que condiciona e incide en las características de rendimiento y en los parámetros de calidad. Se han detectado genes que tienen efecto directo sobre los atributos de la carne (Candek-Potokar et al., 1998; Jeremiah et al., 1999; Wood et al., 1996, Stuczyńska et al., 2018). Muchos de estos genes pueden convertirse en herramientas útiles si son considerados como marcadores moleculares. Entre ellos se encuentra el gen de Halontano (Hal o Ryr1) que, ubicado en el cromosoma 6 porcino, codifica para el receptor de liberación de calcio en el musculo esquelético (Bastos et al., 2001). La mutación no sinónima C1843T produce carnes pse (pale, soft, exudative) pálidas, blandas y exudativas consideradas de menor calidad y de menor valor para procesos industriales (Fujii et al., 1991; Shen et al., 1992, Schilling et al., 2003). Antiguamente, cuando la selección se basaba únicamente en características fenotípicas, el alelo mutado fue ampliamente distribuido por la apariencia hipermusculada que le confería a los animales portadores y al alto contenido magro que estos presentaban. En la actualidad, se lo conoce por su acción perjudicial sobre las características organolépticas de la carne. Además, la misma mutación puntual en el gen es causante del Síndrome de Estrés Porcino (sep) o Síndrome de Hipertermia Maligna, relacionado a una alta tasa de mortalidad por estrés (Otsu et al., 1992, Calvo et al., 1997). Razas como Pietrain, Poland China y Landrace con características magras y con gran desarrollo muscular, implicaban animales portadores del sep. El sep se ha descrito como una enfermedad hereditaria autosómica recesiva (Christian, 1972). Es decir que, cuando los cerdos homocigotas recesivos (tt) son expuestos a situaciones de estrés, durante el manejo pre-sacrificio puede desencadenarse la muerte súbita del animal y la generación de carnes pse (Harrison, 1981). Los animales heterocigotas (Ct) muestran un fenotipo intermedio entre el homocigota para la mutación y el homocigota portador de alelos normales (Silveira et al., 2011). Otra manifestación de las carnes pse es la condición bicolor de los jamones, músculos claros y oscuros, que los desvalorizan para su industrialización (Bastos et al., 2001, Marini et al., 2012).
Posteriormente fueron detectadas varias regiones polimórficas en el genoma porcino y numerosos SNPs (Polimorfismos de Nucleótido Único) de interés para el estudio de calidad de carne, dos de ellos en el gen SOX6. Según Heidt y colaboradores (2013), SOX6 codifica para un factor de transcripción que tiene un alto índice de heredabilidad. La versatilidad de este gen juega un rol importante en la especificación de la fibra lenta durante la diferenciación del músculo esquelético mediante la inhibición de la transcripción de varios genes sarcoméricos (Hagiwara, 2011, Quiat et al., 2011), además se lo asocia con el crecimiento muscular y características de calidad. Uno de estos SNPs (dbSNP ID: rs81358375) está localizado en el intrón 3 de SOX6-like (Entrez Gene ID: 100738152) y el otro (dbSNP ID: rs321666676) está localizado en el exón 7 de SOX6 (Entrez Gene ID: 397173), vecino al extremo 3’ de SOX6 like. Estos SNPs fueron relacionados con el pH, la cra y el color en poblaciones Pietrain y DuPi (Duroc x Pietrain) (Zhang et al., 2015). La fibra muscular es un factor determinante para la calidad de la carne (Klont et al., 1998) y sus propiedades bioquímicas y fisiológicas derivan en sus respuestas divergentes con respecto al estrés previo al sacrificio, la disminución del pH post mortem y la calidad de la carne (Karlsson et al., 1999).
La mayor parte de los músculos post-rigor contienen aproximadamente un 70 % agua, dependiendo primeramente del contenido lipídico y de la madurez fisiológica del músculo (Kauffman, y Warner, 1993). La cra, es la capacidad que tiene la carne de retener el agua cuando es sometida a fuerzas externas, excepto a la gravedad. La cra es causada en primer lugar por la inmovilización de agua de los tejidos en el sistema miofibrilar. El agua más fácil de extraer es el agua extracelular, la que origina la merma por goteo, mediciones de esta agua liberada son usadas como indicador de las propiedades de ligar el agua de las proteínas. La cra y merma por goteo son moderadamente heredables y están correlacionados negativamente una con la otra (Popp et al., 2018). Las medidas de merma por goteo son muy importantes en la industria y también para la apariencia y palatabilidad de la carne fresca. Diferencias en las mermas por goteo son el resultado de un incremento en el metabolismo y una tasa rápida de glucólisis post mortem que da lugar a una disminución muy rápida del pH (por debajo de 5.5-5.8) en la primera hora luego del sacrificio (Guardia et al., 2004). El bajo pH, la elevada temperatura muscular y la acumulación de lactato se traducen en la desnaturalización de las proteínas actina y miosina, la desorganización de las fibras, cambios en la membrana celular, pérdida de la cra (Offer y Cousins, 1992, Hughes et al., 2014) volcado del agua al espacio extramiofibrilar y finalmente la palidez de la carne (Offer y Trinick, 1983). El valor del pH es otro buen indicador de la calidad de carne porcina (Huff-Lonergan y Lonergan, 2005). El pH medido a las 24 horas luego del sacrificio está asociado con la merma por cocción y la dureza en carnes cocidas (Eikelenboom et al., 1996) mientras que los valores de pH a los 45 minutos están asociados con el color: altos valores de pH están relacionados con un bajo valor de la luminosidad de color (*L) (Lindahl et al., 2006).
Dado los antecedentes de SOX6 en la determinación del tipo de fibra muscular este trabajo pretende establecer la incidencia de los polimorfismos de los genes Hal y SOX6 así como también analizar el efecto del genotipo sobre parámetros relacionados a la calidad de la carne porcina en una población del noreste entrerriano.
Materiales y métodos
Obtención y manejo de muestras
Se obtuvo adn de 294 muestras de cerdos del noreste entrerriano. Del total, 239 correspondieron a muestras de pelo de los parentales, las cuales fueron conservadas en oscuridad en bolsas plásticas, a temperatura ambiente hasta el momento de la extracción. Las 55 restantes corresponden a muestras de carne que fueron tomadas de algún miembro de la progenie para llevar a cabo los análisis de calidad. Para obtener estas muestras, el sacrificio se estableció entre el quinto y sexto mes de vida, cuando los animales adquirían un peso entre 90 y 110 kg con una alimentación comercial a base de maíz. Los animales identificados con su correspondiente caravana, fueron transportados en camiones y mantenidos en corrales bajo condiciones de ayuno 12 horas previas a la matanza. Posteriormente fueron sacrificados bajo condiciones industriales, mediante insensibilización eléctrica (1,3 A), izados, desangrados (2 minutos), escaldados y pelados (con agua limpia a 60 °C durante 5 minutos) eviscerados y lavados. Se separaron y marcaron las medias reses destinadas a este estudio y a continuación se pesaron, tipificaron y sellaron de acuerdo a la normativa vigente. Se derivaron a la cámara de oreo, donde quedaron depositadas hasta obtener el resultado de libre triquina. Seguidamente las medias reses fueron enviadas a la cámara de enfriamiento a 0 °C con el fin de llegar a una temperatura de 7 °C para ser despachadas a carnicería. A las 24 horas del sacrificio, se tomó una muestra de músculo de cada animal (Longissimus dorsi) de la región que abarca de la décima a la décimo tercera costilla, realizando cortes de aproximadamente 2 cm de grosor y se las almacenó a temperatura de refrigeración (4 ± 1 °C).
Determinaciones analíticas
pH
Se determinó el pH inicial en el músculo longissimus dorsi, sobre una sección adyacente a la 10ª costilla a los 45 minutos de sacrificado el animal en el predio destinado a la matanza. Posteriormente se realizó una segunda medición a las 24 horas. Las determinaciones se realizaron con el auxilio de un pHmetro portátil (Oakton modelo pH11) equipado con un electrodo de vidrio de penetración (Oakton modelo 35805-18) registrándose un promedio de tres lecturas efectuadas en distintos lugares de la misma sección.
Color
Se determino el color a las 24 horas luego del sacrificio, sobre una sección correspondiente a la 10ª costilla. Las muestras se dejaron oxigenar por 30 minutos y posteriormente se realizaron las determinaciones por triplicado. Se empleó un equipo Minolta CR-700, utilizando el espacio de color cielab (CIE, 1978), iluminante A y ángulo de observador de 10°. Se midieron las coordenadas L* (luminosidad), a* (coordenada rojo-verde) y b* (coordenada amarillo-azul) (AMSA, 2012) (Bertram et al., 2000).
Merma de agua por goteo
Según el método de Honikel (1987), se determinó la pérdida de agua por goteo de una sección correspondiente a la 11ª costilla. Se midió la pérdida de peso del trozo de musculo escurriendo en una bolsa plástica a 4 ºC, a las 24 y 48 horas post mortem. Se expresó en forma porcentual respecto del peso de la muestra fresca.
Mermas de cocción
Previo a la cocción, todas las muestras fueron preservadas bajo congelación por un lapso de dos meses. A las 24 horas de su descongelación en heladera a 4 °C, se procedió a la cocción de las mismas siguiendo la metodología propuesta por el amsa (1995). En un grill George Foreman se colocó una sección correspondiente a la 10ª costilla de aproximadamente 2 cm de grosor. Se colocó un sensor de temperatura en el centro geométrico de cada bife, los valores fueron registrados mediante Yokogawa, mod. DX106-1-2. Cuando la temperatura alcanzó los 71 °C la muestra fue retirada del grill y se dejó enfriar a temperatura ambiente. A continuación, se pesó para determinar las mermas producidas por la cocción. Las mismas fueron expresadas en porcentaje respecto del peso de los bifes previo a la cocción.
Terneza
Se evaluó la fuerza máxima de corte con el equipo Stable Micro Systems TA - XT2i utilizando la célula Warner-Bratzler, sobre cilindros de 1,27 cm de diámetro cortados con la fibra paralela al eje longitudinal. Se determinó fuerza máxima de cizalla en kgf (amsa, 1995) y se registró el valor medio de un mínimo de 8 cilindros extraídos de cada muestra. La muestra corresponde a una sección de la 10ª costilla cocida 24 horas antes, almacenada en una bolsa de polipropileno a la temperatura de refrigeración de 4 °C.
Humedad
La humedad se determinó por desecación en estufa a 125 °C durante 4 horas, de acuerdo con el método 950.46 descrito por la aoac (2007). Se registró el valor medio de 5 réplicas para cada muestra, posteriormente se expresaron los resultados como porcentaje con respecto al total de la muestra analizada.
Técnicas Moleculares
Extracción de ADN a partir de muestra de pelo
La obtención de adn genómico se realizó a partir de bulbos de pelos extraídos mediante el método del bromuro de cetil-trimetil amonio (ctab) (Murray y Thompson 1980, Sambrook et al., 1994). Se incubaron 15 bulbos en buffer te, 10 % sds y proteninasa K (1mg/ml) por 15 minutos a 37 °C. Luego, se adicionó NaCl 5M y ctab (0,7M NaCl, 10 % ctab, Genbiotech) y se incubó a 65 °C por 10 minutos. Posteriormente se transfirió la solución a un nuevo tubo evitando acarrear restos de pelo y se adicionó cloroformo: alcohol isoamílico en una proporción 24:1, se centrifugó a 13000 rpm por 12 minutos y la fase acuosa se transfirió a un nuevo tubo. Para precipitar el adn se adicionó isopropanol frío, se mezcló por inversión y se centrifugó a 13000 rpm 6 minutos. Finalmente, se realizaron dos lavados del pellet con de etanol 70 %, se dejó secar a temperatura ambiente y se resuspendió en 15 μl de buffer te.
Extracción de ADN a partir de muestras de músculo
Se pesaron entre 0,02 y 0,03g de carne procesada previamente con mortero, luego cada muestra se resuspendió en buffer te y se utilizó el mismo protocolo de extracción con ctab que es utilizado para las muestras de pelo.
Cuantificación y evaluación del adn obtenido
La concentración de la muestra de adn se calculó teniendo en cuenta el valor de absorbancia obtenido a una longitud de onda de 260 nm. Mientras que la relación de absorbancias A260/280 se utilizó para evaluar la pureza de las muestras.
Amplificación de adn mediante pcr
Para cada muestra de adn, la mezcla de pcr estuvo compuesta por dNTPs 1 mM, primers Forward y Reverse, agua, green buffer 10X y adn polimerasa más el adn purificado en un volumen final de 25 μl. Los primers utilizados figuran en la Tabla 1. La amplificación por pcr se realizó en un termociclador pcr convencional esco aeris con el siguiente programa de ciclado: desnaturalización inicial a 94 ºC por 5 min.; 38 ciclos de 94 ºC por 30 seg., temperatura de annelling específica para cada par de primers, por 30 seg. y 72 ºC por 30 seg., y una extensión final a72 ºC por 7 min. Una alícuota de 5ul del producto de pcr será sometida a electroforesis en gel de agarosa al 1 % con 0,1 μg/ml de bromuro de etidio, visualizada con transiluminador uv y fotografiada usando una cámara digital Kodak EasyShare Z7590.

Genotificación mediante polimorfismos de longitud de fragmentos de restricción (rflp)
Una alícuota de 10µl del producto de pcr fue digerida con enzimas de restricción (Tabla 1), de acuerdo al protocolo del fabricante. Luego, esta digestión fue sometida a electroforesis en gel de agarosa al 3 % con 0,1 μg/ml de Bromuro de Etidio visualizado con Transiluminador uv y fotografiada.
Análisis estadísticos
El efecto de las distintas variantes genotípicas sobre los atributos de la carne se evaluó mediante un análisis de anova simple para cada polimorfismo de hal y SOX6. Considerando las variantes genotípicas como un factor fijo con cuatro niveles para hal y SOX6a (ccaa, ccag, ctaa y ctag) y para hal y SOX6b (ccgc, ccgg, ctgc y ctgg). Para comparar las medias se utilizó el test lsd con un α: 0,05. El análisis de los datos se efectuó mediante el programa Statgraphic centurion xv (StatPoint Tech, Inc., Warrenton, va, usa). Los genotipos ctgg para hal y SOX6a y cccc para hal y SOX6b no fueron considerados para el análisis de la varianza por encontrarse representados por una única muestra.
Las frecuencia alélicas y genotípicas se calcularon según la siguiente fórmula: frecuencia genotipo A + ½ frecuencia genotipo ab.
Resultados y discusión
Los resultados obtenidos en este trabajo permiten brindar datos fehacientes a los productores para orientarlos en la aplicación de la selección asistida por marcadores moleculares con el objetivo de disminuir la incidencia de alelos perjudiciales y aumentar la de aquellos que resulten beneficiosos para llegar al mercado con carnes de mejor calidad.
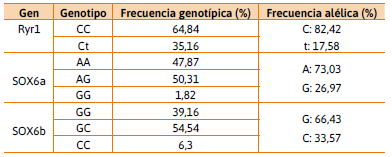
Frecuencias alélicas y genotípicas
Se caracterizó la población en estudio de acuerdo a las frecuencias alélicas y genotípicas para los genes Halotano y SOX6. En la Tabla 2, se representan las frecuencias observadas para las 294 muestras porcinas analizadas. Para el gen Ryr1, el alelo normal C se encuentra representado por el 82,42 % y el alelo mutado t, relacionado con carnes PSE, por el 17.58 % siendo las frecuencias genotípicas 64,84 % para cc y 35,16 % para el genotipo Ct. Aunque no se han detectado animales con el genotipo tt, la incidencia del alelo perjudicial se evidencia en heterocigosis. Estos datos demuestran una menor frecuencia del alelo t con respecto a los resultados observados por el grupo de Marini y colaboradores (2012) (cc 65 %, Ct 30,8 % y tt 4,2 %) en poblaciones de animales híbridos en provincias mediterráneas (Córdoba, Santa Fe, Chaco y Tucumán) y aún menor a las observadas en granjas comerciales del sur de Brasil (Bastos et al., 2000; Oliveira Vargas et al., 2002; Band et al., 2005, Silveira et al., 2011). Dado que en el presente trabajo no se discrimina el origen genético de los animales (criollos o líneas híbridas comerciales) es destacable la baja frecuencia del alelo t en la población analizada. Para el gen SOX6 se estudiaron dos SNPs SOX6a (a/g) y SOX6b (c/g), para el polimorfismo SOX6a se observaron las siguientes frecuencias genotípicas: AA 47,87 %, AG 50,31 % y gg 1,82 %, que evidencian una alta frecuencia del alelo A, siendo ésta mayor a la reportada en 2015 (17, 47 y 35 % para Pietrain 36, 48 y 19 % para DuPi); mientras que para el polimorfismo SOX6b se observa una elevada frecuencia del alelo G distribuyéndose en los siguiente genotipos: gg 39,16 %, gc 54,54 %.; y si bien el genotipo cc no fue observado por Zhang y colaboradores (2015), en esta población del noreste entrerriano se encuentra representado en un 6,3 %. Estos resultados constituyen un segundo acercamiento a un análisis poblacional de los polimorfismos asociados al factor de transcripción SOX6 luego del trabajo publicado por Zhang y colaboradores (2015).
Efecto de los polimorfismos de Halotano (Ryr1) y SOX6a sobre los parámetros de calidad de carne
De las 55 muestras analizadas para calidad de carne, 20 resultaron portadoras de la mutación en el gen de Ryr1 (Ct), mientras que las restantes 35 corresponden al genotipo cc, libres de la mutación. Al realizar las diferentes combinaciones con los polimorfismos de Hal y SOX6a, se obtienen los siguientes genotipos con sus correspondientes frecuencias: ccaa 24,46 %, ccag 38,18 %, ctaa 20 %, ctag 14,54 % y ctgg 1,82 %. No se encontraron animales ccgg y a fines del análisis estadístico, el genotipo ctgg fue descartado por estar representado por una única muestra. Los valores obtenidos en las diferentes determinaciones realizadas se expresan como media ± ds en la Tabla 3.
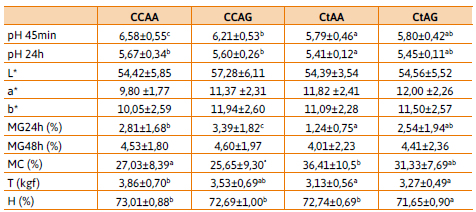
Se observaron diferencias significativas (p < 0,05) entre los valores medios de pH a los 45 minutos y a las 24 horas post mortem para los diferentes genotipos (Tabla 3). Ambas observaciones evidencian valores mayores de pH para el genotipo ccaa seguido por ccag y los menores para los genotipos portadores de la mutación para Halotano (ctaa y ctag), datos ampliamente concordantes con la literatura (Silveira et al., 2011). En animales libres de Hal, aquellos cerdos que presentaron el genotipo aa tienen un pH más elevado coincidiendo con los resultados obtenidos por Zhang y colaboradores (2015).
Las mermas son usadas para estimar la cra y resultan inversamente proporcionales a esta (Popp et al., 2018). En este sentido se observaron diferencias significativas (p < 0,05) en las mermas por goteo a las 24 horas, donde los genotipos heterocigotas para SOX6a (ccag y ctag) presentan las mermas más altas. Si bien no se observaron diferencias significativas en las mermas por goteo a las 48 horas se evidencia una tendencia similar a la observada a las 24 hrs.
También se manifiestan diferencias significativas (p < 0,05) entre los valores medios de terneza y los diferentes genotipos, siendo el genotipo ccaa el que presenta mayor fuerza al corte (menor terneza) seguido por ccag, ambos homocigotas normales para Halotano (Tabla 3). Los patrones post mortem de conversión de músculo a carne dependen del tipo de fibra predominante en el músculo. En la literatura se destaca el rol de SOX6 como modulador de la diferenciación de las fibras, elevados niveles de este factor de transcripción inhiben la actividad de los genes que codifican las fibras tipo I (Hagiwara et al., 2005; Hagiwara et al., 2007; von Hofsten et al., 2008), por lo que las fibras tipo II serían predominantes. Otros factores podrían afectar el parámetro terneza como cambios en el metabolismo energético de las fibras pre y post mortem, la expresión diferencial de los factores de transcripción fibroespecíficos, así como también la expresión de reguladores de la expresión de SOX6 como miR-208b y miR-499-5p (Zhang et al., 2015). Sería necesario realizar estudios de expresión del factor SOX6 en relación a los fenotipos observados. La modificación de las secuencias de unión de estos podrían explicar el fenotipo observado para ccaa (Zhang, et al., 2015).
Por otro lado, se encuentran diferencias significativas (p < 0,05) entre los valores medios de humedad (h) para los diferentes genotipos observados. ccaa presenta las mayores mediciones para humedad en carne fresca que se corresponden con el elevado pH observado a los 45 minutos post mortem. Todos los genotipos resultaron en valores similares para los parámetros de color L*, a* y b* por lo que este SNP no estaría relacionado a este atributo.
Los resultados muestran el alelo perjudicial de Hal se encuentra asociado con los valores de pH, humedad y terneza y que a su vez el alelo A de SOX6a podría presentar efecto un positivo sobre el pH y cra. El genotipo ccaa podría resultar el más beneficioso para la generación de carne fresca de calidad.
Efecto de los polimorfismos de Halotano (Ryr1) y SOX6b sobre los parámetros de calidad de carne
Para este polimorfismo también se realizaron las combinaciones con el gen Hal y se analizaron las frecuencias para los genotipos observados en la población. Obteniéndose 25,45 % ccgc, 36,36 % ccgg, 12,72 % ctgc, 23,64 % ctgg y 1,83 % cccc, este último fue descartado del análisis estadístico por estar representado solo por una única muestra. En la Tabla 4 se observan los datos obtenidos para las determinaciones de la calidad de carne para cada genotipo.
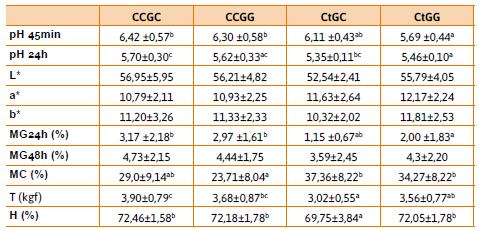
Se observaron diferencias significativas (p < 0,05) entre los valores medios de pH a los 45 minutos y a las 24 horas post mortem para los diferentes genotipos. Las muestras portadores del alelo t (ctgc y ctgg) presentan los valores más bajos de pH, concordando con Silveira y colaboradores (2011) quienes observaron que animales heterocigotas (Ct) manifestaban un fenotipo intermedio entre el homocigota para la mutación y el homocigota portador de alelos normales. A su vez, el genotipo CtGG presenta los valores más bajos de pH 45 sugiriendo que el alelo G de SOX6b estaría asociado con un descenso del pH, mostrando un efecto aditivo al alelo t.
Los resultados arrojan también diferencias significativas (p < 0,05) para los valores medios de mg a las 24 horas, observándose una mayor merma en los genotipos ccgg y ccgc, lo que podría sugerir un efecto del alelo g sobre este parámetro. Esta mayor merma implicaría una mayor resistencia al corte, tal y como se ve reflejado en los valores obtenidos (3,90±0,79). La diferencia de pH observada para los individuos portadores de alelo perjudicial de Hal, se ve reflejada también en las mc donde los genotipos ctcg y ctgg presentan valores más elevados que su contraparte homocigota (p< 0,05). En adición el genotipo ctgc resulta el de menor contenido de humedad en carne fresca, menores valores de pH y mermas por goteo a las 24 horas.
En poblaciones Piertrain y DuPi se relacionó el alelo G con el color (Zhang et al., 2015), los valores obtenidos para los parámetros de color L*, a* y b* resultan similares entre los diferentes genotipos para la población aquí analizada, donde el alelo g presenta relación con el valor de pH.
Conclusiones
La selección asistida por marcadores nucleicos constituye una herramienta útil en la producción pecuaria, permite elegir los animales con características deseables tomando en cuenta la información obtenida del adn del individuo, reduce la necesidad de colectar información fenotípica, genera un aumento en la precisión al seleccionar los animales y disminuye el intervalo generacional. A través del uso de esta herramienta, en el presente trabajo se logró caracterizar una población del noreste entrerriano en cuanto a los polimorfismos de los genes Hal y SOX6 relacionados a la calidad de la carne. De los resultados obtenidos se desprende que aun existe una alta incidencia (17,58 %) del alelo perjudicial de Halotano, alelo t. Si bien este se encuentra en heterocigosis, este genotipo puede conducir no solo a carnes de baja calidad sino también a pérdidas económicas para los productores al generar descendencia susceptible al Síndrome del Stress Porcino. En cuanto a los polimorfismos asociados al gen SOX6 sugieren una relación con los parámetros de pH y cra. Particularmente para SOXa, el alelo A representado en un 73,03 %, podría presentar un efecto sobre el pH y la cra resultando beneficioso para la calidad de carne. Con respecto al polimorfismo SOX6b el alelo g, presente en el 66,43 % de la población estudiada podría estar vinculado con un descenso del pH. Siendo los genotipos portadores de los alelos c para Halotano, A para SOX6a y c para SOX6b los genotipos ideales para carne fresca.
Referencias bibliográficas
American Meat Science Association (amsa). (2012) Meat Color Measurement Guidelines Illinois: American Meat Science Association - National Livestock and Meat Board. Illinois.
American Meat Science Association (amsa). (1995). Research guidelines for cookery, sensory evaluation and instrumental tenderness measurements of fresh meat. Illinois: American Meat Science Association - National Livestock and Meat Board. 48p.
AOAC (2007). Official methods of analysis of aoac international. 18th ed. Washington: Association of Official Analytical Chemists.
Band, G.O.; Guimarães, S.E.F.; Lopes, P.S.; Schierholt, A.S.; Moraes Silva, K.; Vieira Pires, A.Benevenuto Júnior, A., Miranda Gomide, L.A. (2005) Relationship between the Porcine Stress Syndrome gene and pork quality traits of F2 pigs resulting from divergent crosses. Genet Mol Biol, 28(1):88-89.
Bastos, R.G.; Federizzi, J.; Deschamps, J.C.; Cardellino, R.A.; Dellagostin, O.A. (2001). Efeito do Gene do Estresse Suíno sobre Características de Quantidade e Qualidade de Carcaça. Rev Bras Zootec, 30(1):37-40.
Bertram, H.C.; Petersen, J.S., Andersen, H.J. (2000).Relationship between rn (-) genotype and drip loss in meat from Danish pigs. Meat Sci, 56(1):49-55.
Calvo, J.H.; Osta, R.; Garcia-Muro, E., Zaragoza, P. (1997). Síndrome de estrés porcino: aplicación y ventajas de la pcr para su diagnóstico. Med. Vet,14. 2: 110-113.
Candek-Potokar, M.; Zlender, B.; Lefaucheur, L., Bonneau, M. (1998). Effects of age and/or weight at slaughter on longissimus dorsi muscle: Biochemical traits and sensory quality in pigs. Meat Sci, 48:287-300.
Chan, W.K.M.; Faustman, C.; Decker, E.A. (1997). Oxymyoglobin oxidation as affected by oxidation products of phosphatidylcholine liposomes. J Food Sci, 62: 709-712.
Chan, W.K.; Faustman, C.; Velasquez-Pereira, J.; McDowell, L.R., Batra, T.R. (1998). Effects of alpha-tocopherol on metmyoglobin formation and reduction in beef from cattle fed soybean or cottonseed meal diets. J Anim Sci, 76: 1421-6.
Christian, L.L. (1972). A review of the role of genetics in animal estress susceptibility and meat quality. Proceedings of the quality symposium, Wisconsin University, Madison 91 -115.
CIE (1978). Supplement 2 to cie Publication 15 (1971). Recommendations on uniform color spaces, color difference equations, psychometric color terms. Paris: Bureau Central de la cie.
Faustman, C.; Chan, W.K.; Schaefer, D.M., Havens, A. (1998). Beef Color Update: The Role for Vitamin E. J Anim Sci, 76:1019-26.
Fujii, J.; Otsu, K.; Zorzato, F.; de Leon, S.; Khanna, V.K:; Weiler, J.E.; O’Brien, P.J., MacLennan, D.H. (1991). Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia. Science, 26; 253(5018):448-51.
Guárdia, M.D.; Estany, J.; Balasch, S.; Oliver, M.A., Gispert, M. (2004). Risk assessment of pse condition due to pre-slaughter conditions and RYR1 gene in pigs. Meat Science, 67:471- 478.
Hagiwara, N. (2011). Sox6, jack of all trades: a versatile regulatory protein in vertebrate development. Dev Dyn 240:1311-1321.
Hagiwara, N.; Yeh, M., Liu, A. (2007). Sox6 is required for normal fiber type differentiation of fetal skeletal muscle in mice. Dev Dyn 236:2062-2076.
Harrison, G. (1981). The malignant hiperthermia porcine. Int Anesthesiol Clin,17 (4): 19-47.
Honikel (1987). Influence of Chilling on Meat Quality Attributes of Fast Glycolysing Pork Muscles.Evaluation and Control of Meat Quality in Pigs. Current Topics in Veterinary Medicine and Animal Science, 38: 273-283.
Huff-Lonergan, E., Lonergan, S.M. (2005). Mechanisms of water-holding capacity of meat: The role of postmortem biochemical and structural changes. Meat Sci,71:194-204.
Hughes, J.M.; Oiseth, S.K.; Purslow, P.P., Warner, R.D. (2014). A structural approach to understanding the interactions between colour, water-holding capacity and tenderness. Meat Sci, 98:520-532.
Jeremiah, L.E.; Gibson, J.P.; Gibson, L.L.; Ball, R.O.; Aker, C., Fortin, A. (1999). The influence of breed, gender, and PSS (Halothane) genotype on meat quality, cooking loss, and palatability of pork. Food Research International, 32:59-71.
Karlsson, A.; Enfalt, A.C.; Essen-Gustavsson, B.; Lundstrom, K.; Rydhmer, L., Stern, S. (1993). Muscle histochemical and biochemical properties in relation to meat quality during selection for increased lean tissue growth rate in pigs. J Anim Sci, 71:930-938.
Kauffman, R.G., Warner R.D. (1993). Evaluating pork carcasses for composition and quality. In G.R. Hollis, Growth of the pigs. Wallingford, UK: CAB International, 141-166.
Klont, R.E.; Brocks, L., Eikelenboom, G. (1998). Muscle fibre type and meat quality. Meat Science 49, Supplement 1:S219-S229.
Lindahl, G.; Enfält, A.C.; von Seth, G.; Josell, A.; Hedebro-Velander, I.; Andersen, H.J.; Braunschweig, M.; Andersson, L., Lundström, K. (2004). A second mutant allele (V199I) at the PRKAG3 (rn) locus- I. Effect on technological meat quality of pork loin. Meat Sci, 66 (3):609- 19.
Marini, S.J.; Vanzetti, L.S.; Borelli, V.; Villareal, A.; Denegri, G.; Cottura, G.; Panichelli, D.; Silva, P.; Campagna, D., Brunori J., Spiner N.; Franco, R. (2012). RYR1 gene variability and effect on meat pH in Argentinean hybrids swines. InVet.,14(1).
Murray, M.G. y Thompson, W. F. (1980). Rapid isolation of high molecular wieght plant dna. Nucleic Acids Res, 8(19) 4321-4325.
Offer, G., Cousins, T. (1992). The mechanism of drip production: Formation of two compartments of extracellular space in muscle Post mortem. J Sci Food Agric, 58:107116.
Oliveira-Vargas-Culau, P.; López, J.; Rubensam, J.M.; Félix-Lopes, R.F., Nicolalewsky, S. (2002). Influencia do gene Halotano sobre a qualidade da carne súina. R. Bras. Zootec, 31(2): 964-96.
Otsu, K.; Phillips, M.S.; Khanna, V.K.; Leon, S., Mac Lennan, D.H. (1992). Refinement of diagnostic assays for a probable causal mutation of porcine and human malignant hyperthermia. Genomics, 13: 835.
Popp, J.; Sharifi, A. R.; Janisch, S., & Krischek, C. (2018). Two Frequently Used pH Determination Methods Showed Clearly Different pH Slopes During Early Post Mortem Ageing of Pork. Food Analytical Methods, 11(6):1773–1779.
Quiat, D.; Voelker, K.A.; Pei, J.; Grishin, N.V.; Grange, R.W.; Bassel-Duby, R., Olson, E.N. (2011). Concerted regulation of myofiber-specific gene expression and muscle performance by the transcriptional repressor Sox6. Proc Natl Acad Sci U S A, 108:10196-10201.
Sambrook, J.; Fritsch, E.F.; Maniatis, T. (1994). Molecular Cloning: A laboratory manual. 2nd ed. New York: Cold Spring Harbor. 3v.
Schilling, M.W.; Mink, L.E.; Gochenour, P.S.; Marriott, N.G., Alvarado, C.Z. (2003). Utilization of pork collagen for functionality improvement of boneless cured ham manufactured from pale, soft, and exudative pork. Meat Sci, 65(1):547-53.
Shen, H.; Lahucky, R.; Kovac, L.; O’ Brien, P.J. (1992). Comparison of Hal gene status with PNMR-determined muscle metabolites and with Ca sequestration activity of anoxia -challenged muscle from pigs homozygous and heterozigous for porcine stress syndrome. Pig News and Information, 13: 105-109.
Silveira, A.C.P.; Freitas, A.S.M.; Cesar, R.C.; Antunes, E.C.; Guimaraes, D.F.A.; Batista, D., Torido, L.C. (2011). Influence of the halotane gene (HAL) on pork quality in two comercial crossbreeds. Genetics and Molecular Research, 10 (3): 1479-1489FA.
Stuczyńska A, Piórkowska K, Tyra M, Żukowski K. (2018). The effect of qtl-rich region polymorphisms identified by targeted dna-seq on pig production traits. Mol Biol Rep, 45(3):361- 371.
Suman, S.P., Faustman, C., Stamer, S.L., Liebler, D.C. (2007). Proteomics of lipid oxidation- induced oxidation of porcine and bovine oxymyoglobins. Proteomics, 7: 628-640.
USDA. United State Department of Agriculture. United States Standars for Grades of Carcass Beef. 1997.
Vestergaard, M., Oksberg, N., Henckel, P. (2000). Influence of feeding intensity, grazing and finishing feeding on muscle fibre characteristics and meat colour of semitendinosus, longissimus dorsi and supraspinatus muscles of young bulls. Meat Science, 54: 177-185.
Von Hofsten, J.; Elworthy, S.; Gilchrist, M.J.; Smith, J.C.; Wardle, F.C., Ingham, P.W. (2008). Prdm1- and Sox6-mediated transcriptional repression specifies muscle fibre type in the zebrafish embryo. EMBO Rep, 9:683-689.
Wood, J.D.; Brown, S.N.; Nute, G.R.; Whittington, F.M.; Perry, A.M.; Johnson, S.P., Enser, M. (1996). Effects of breed, feed level and conditioning time on the tenderness of pork. Meat Sci, 44:105-112.
Zhang, R.; Grobe-Brinkhaus, C.; Heidt, H.; Jasim Uddin, M.; UlasCinar, M.; Tesfaye, D.; Tholen, D.; Looft, C.; Schellander, K.; Neuhoff, C. (2015). Polymorphisms and expression analysis of SO X- 6 in relation to porcine growth, carcass, and meat quality traits. Meat Science, 107:26– 32.
Notas de autor
lagadarim@fcal.uner.edu.ar